|
|
Эволюционная роль половых хромосом. (Новая концепция)
В.А. Геодакян
(Опубликовано в журнале "Генетика", 1998, т. 34, №
8, с. 1171-1184)
“Представляется, что человеческий разум должен свободно
строить формы, прежде чем подтвердится их реальное существование...
Из голой эмпирии не может расцветать познание. Такой расцвет возможен
только из сравнения придумываемого и наблюдаемого”.
Альберт Эйнштейн2
“Одной из основных задач теоретического исследования
в любой области знания является установление такой точки зрения,
с которой объект исследования проявляется с максимальной простотой”.
Виллард Гиббс3
Половая дифференциация позволяет эволюционные новации
проверять в мужском геноме, прежде чем передавать их в женский.
Это возможно при дихронной (асинхронной) эволюции, когда эволюция
мужского пола предшествует эволюции женского4.
Значит, наряду с локализованными в аутосомах общими генами стабильных
признаков, должны существовать сугубо мужские, уже попавшие в мужской
геном, но еще не попавшие в женский, и сугубо женские, уже утраченные
мужским полом, но еще остающиеся у женского, составляющие временный
эволюционный ГПД5.
По общим же генам не может быть ГПД, они образуют только постоянный
ФПД (гормональный). Такая трактовка ГПД, как следствия полового
дихронизма, позволяет выявить эволюционные роли ПХ и предложить
для них новую концепцию, согласно которой Y-хр - “ворота” для экологической
информации в геном, место “рождения” и апробации новых генов, инициатор,
акселератор и регулятор ГПД, Х-хр гетерогаметного пола - “транспортная”,
переносит новые гены из Y-хр в А-, является стабилизатором, релаксатором
и ликвидатором ГПД, а также хранилищем отработавших, ожидающих элиминацию
генов. Концепция дает новый подход к пониманию многих явлений: появлению,
локализации и переходам генов по хромосомам и между ними, инактивации
хромосом, мобильных генов, “вспышек” мутаций, инсерционного мутагенеза,
связи Y-хр со стрессом, ретровирусами и др. В частности, позволяет
объяснить куда и зачем “прыгают” гены, почему транспозиции мобильных
элементов зависят от экологического стресса, почему мутируют одновременно
разные гены и т.д.6
ВВЕДЕНИЕ
Со времени открытия ПХ7,
считается, что их основная роль - определение пола и обеспечение
СП 1:1. А так ли это? Конечно, ПХ делают и то и другое, но можно
ли на этом основании считать, что в этом их главное функциональное
значение? Ведь для определения пола вполне достаточно одного А-гена-триггера,
а СП 1:1 автоматически получается при скрещивании рецессивной гомозиготы
с гетерозиготой. Ведь пол существовал и до появления ПХ и многие
современные РП виды не имеют ПХ. Тогда не ясен эволюционный смысл
(роль, целесообразность, назначение) аутосомно-гоносомной ДИФ. Какой
принцип лежит в ее основе? Что дают разные алгоритмы поведения хромосом?
Почему А- передаются от родителей к потомству чисто стохастически,
случайно, а ПХ - по особым маршрутам: Y-хр от отца - только сыну,
Х-хр - только дочери? Какие гены локализованы в А-, какие - в Х-хр,
какие - в Y-? Как можно объяснить особенности конъюгации,
кроссинговера или конденсации ПХ в зависимости от пола и типа гаметности?
Мы много знаем о работе генов в хромосомах в онтогенезе, но
почти ничего не знаем об их “образе жизни” в геноме в филогенезе.
Ведут ли они себя “оседло”: “рождаются”, “живут”, работают и “умирают”
в одной хромосоме или - “кочуют”: разные фазы своего филогенеза
проводят в разных хромосомах? Существует ли закономерный “маршрут”
генов по хромосомам? Если да, то какой?
Накопилось много загадок, противоречий и просто новых
данных, которых классическая теория ПХ не может объяснить. Скажем,
трактовка телец Барра как компенсации дозы Х-генов. Если конденсация
действительно была бы для этого, то тельца Барра должны были в норме
образоваться всегда у гомогаметного пола. Однако у птиц так же,
как у млекопитающих, конденсируется Х-хр самок (причем единственная!).
Какая же это компенсация дозы? У птиц же отсутствует конъюгация
ПХ. Почему-то репликация ДНК одной Х-хр у гомогаметного пола и Y-
хр происходит после окончания репликации аутосом8,
и т.д.
Очень загадочна Y-хромосома. Это самая вариабельная,
особенно по длине, хромосома генома, богатая нуклеотидными повторами
и гетерохроматином у животных, и эухроматином и разбросанными
по всем хромосомам повторами - у растений9.
У человека она генетически почти пустая (если не считать генов волосатости
ушей и перепонок между пальцами ног). У других видов может содержать
много активных генов. Например, у дрозофилы известно много генов,
локализованных внутри Y-гетерохроматина. У гуппи еще в 20-30гг были
открыты свыше 30 Y-генов окраски самцов (и только один А- ген!),
часть из них участвует в неравном кроссинговере с Х-хр, при этом,
переход Y–Х в 4 раза чаще, чем обратный10.
На стрекозах было показано, что форма ХY эволюционно более
поздняя, чем Х0. Однако есть и противоположная точка зрения, согласно
которой, ПХ произошли из обычной пары А-, несущей определяющие пол
гены, поэтому у одних видов (более примитивных) Y-хр такая же по
размерам как Х-хр, конъюгирует с ней полностью или частично и участвует
в кроссинговере. У других видов (более прогрессивных) - она маленькая,
с Х-хр соединяется конец в конец, без кроссинговера. В процессе
эволюции Y-хр почему-то теряет свои активные гены, деградирует и
исчезает. Поэтому форма ХY предшествует Х011.
Этот вопрос остается пока открытым. Только в одном трудно согласиться
с автором, что ярко-красное пятно самцов гуппи раньше было
и у самок, которые его потом утратили. Думаю, оно никогда не было
у самок. Были сообщения о более крупной Y-хр у разных этнических,
или социальных групп, о большой дисперсии ее у грызунов, обитающих
в зонах повышенной сейсмической активности12,
о связи с ретровирусами13,
с новыми мутациями14,
и т.д.
Создается впечатление, что с ПХ, и прежде всего с Y-хр,
мы чего-то существенного не понимаем. А не понимаем главного - для
чего ПХ? Что они делают? Каково их назначение, функциональная роль
и адаптивное значение? В чем их эволюционный смысл, логика? В настоящей
статье предлагается новая концепция ПХ, отвечающая на эти
вопросы, модель, основанная на идее дихронной эволюции. Но прежде,
необходимо ответить на другой, более фундаментальный, вопрос - “для
чего пол?”
ПРОБЛЕМА ПОЛА
На протяжении полутора веков пол остается центральной
проблемой эволюционной биологии. Ею занимались крупнейшие биологи
19-20 вв. - Дарвин, Уоллес, Вейсман, Гольдшмидт, Фишер, Мёллер.
И несмотря на это современные авторитеты продолжают писать в связи
с полом о “кризисе” в эволюционной биологии. За последние 20 лет
проблема пола переживает очередной ренессанс. Появилась дюжина книг,
в заглавии которых фигурируют два слова: “пол” и “эволюция”15.
Одна из них начинается с фразы: “Преобладание полового размножения
у высших растений и животных несовместимо с современной эволюционной
теорией”16. В другой
- читаем: “Мы не имеем удовлетворительного объяснения тому, как
возник и как сохраняется пол”17.
В третьей монографии, посвященной эволюции и генетике пола, автор
пишет: “Проблема пола - главный вызов современной теории эволюции...
Царица проблем... Интуиции Дарвина и Менделя, осветившие так
много загадок, не смогли справиться с центральной загадкой
полового размножения.”18.
Еще один крупнейший авторитет по проблеме пола пишет: “Довольно
поразительно, но ученые не могут убедительно сказать, зачем существует
пол.”19. На эту
тему появляется много статей и обзоров. По крайней мере, два ведущих
генетических журнала посвятили этой проблеме в 1993-94 гг.
специальные выпуски20.
Все это говорит о том, что центральная проблема эволюционной биологии
и генетики - проблема пола - за рубежом остается все еще нерешенной,
и на главный вопрос: для чего пол, какое имеет адаптивное значение,
ответа пока нет21.
То, что у эволюционной теории в вопросах пола концы
не сходятся с концами мне стало ясно в начале 60 гг. Впервые эвристическое
решение проблемы пола было опубликовано мной в 1965 г. в небиологическом
журнале22.
В чем суть проблемы
и почему ее не могут решить?
Самой фундаментальной программой жизни является программа
репродукции, лежащая в основе таких биологических явлений, как репликация,
редупликация, БП размножение. Она является главным критерием, отличающим
живые системы от неживых. Основной источник разнообразия при этой
программе - мутации. Вторая по важности программа жизни -
программа рекомбинации, которая лежит в основе таких биологических
явлений как кроссинговер, оплодотворение или сингамия. Создав
новый - внутренний, автономный от среды, источник разнообразия,
РЕК кардинальным образом решила проблему разнообразия. На ее основе
возник половой процесс. Третьей по важности программой можно
считать программу дифференциации, лежащую в основе явлений мейоза,
половой и других ДИФ. В результате этой программы появились РП формы,
касты у общественных насекомых, карликовые самцы у некоторых рыб
и другие ДИФ. В процессе эволюции эти программы, и все биологические
явления на их основе, возникли именно в таком порядке, отражающем
конститутивно-факультативные отношения между ними, когда предшествующие,
более основополагающие обязательны для существования последующих,
а наличие вторых - факультативно, не обязательно для первых.
Понятие пола включает два фундаментальных явления: половой
процесс (слияние генетической информации двух особей) и половую
дифференциацию (разделение этой информации на две части). В зависимости
от присутствия (+) или отсутствия (-) этих явлений, множество существующих
способов размножения можно разделить на три основные формы: (-,
-) - БП, (+, -) - ГФ и (+,+) - РП (табл.1).
Табл. 1. Характеристики трех основных способов
размножения
|
Способ размно-
жения
|
Программы (биол. явление)
|
Эффективность
|
Источн.
разнообразия
|
|
РЕП
(размно-жение)
|
РЕК
(половой процесс)
|
ДИФ
(половая
диффер.)
|
Количес-твенная
|
Ассорта-
тивная
|
Качест-
венная
|
|
БП
|
+
|
-
|
-
|
max
|
mid
|
min
|
M*
|
|
ГФ
|
+
|
+
|
-
|
mid
|
max
|
mid
|
M + P*
|
|
РП
|
+
|
+
|
+
|
min
|
mid
|
max
|
M+P+Д*
|
* М - мутации, Р - рекомбинации, Д - дифференциация.
Половой процесс и половая ДИФ явления разные и, по своей
сути, диаметрально противоположные. П процесс создает (повышает)
разнообразие генотипов и в этом его эволюционное предназначение.
П ДИФ, наоборот, снижает его вдвое и в чем её эволюционная роль
- никто не может объяснить. Так, в бесполой популяции, состоящей
из N особей, максимальное, теоретичеески возможное, разнообразие
генотипов потомства равно N, в том случае, если генотипы всех родителей
разные. Так как потомство каждой бесполой особи - клон с одинаковым
генотипом, то разнообразие потомства, s, может быть только меньше
N.
С появлением П процесса возможное разнообразие потомства
возводится в квадрат численности. При этом у ГФ каждая из N особей
может скрещиваться с N-1 особью (кроме себя), но поскольку
особь №1 с№2то же самое, что №2 с №1 (т.е.
нет реципрокного эффекта), то при N >>1, разнообразие: s =
N(N-1)/ 2 » N2/2 (при наличии реципрокных
различий s » N2). У РП форм, появление П ДИФ, наложив
запрет на однополые комбинации (мм, жж), минимум вдвое уменьшает,
возможное у ГФ разнообразие: s = N/2 ´ N/2 = N2/4
(каждая самка с каждым самцом, при одинаковой их численности N/2).
Разнообразие потомства в РП популяции зависит еще от СП в родительском
поколении: оно максимально при СП= 1:1, и уменьшается при
любом отклонении от этого значения.
Значит, при одинаковой численности N, максимально возможное
разнообразие потомства БП, ГФ и РП популяций относятся друг к другу
соответственно, как: N : N2/2 : N2/4,
т.е. при переходе от ГФ к РП утрачивается минимум половина разнообразия!
Тогда, совершенно непонятно - что дает ДИФ, если она ухудшает вдвое
основное достижение П размножения? Почему все прогрессивные в эволюционном
плане виды животных: млекопитающие, птицы, насекомые, и растений
(двудомные) - РП, в то время, как явные преимущества количественной
эффективности и простоты у БП, а разнообразия потомства - у ГФ?
В этом суть загадки пола, которую не могут решить. Это объясняется
прежде всего тем, что нет четкого понимания, что П процесс и П ДИФ
противоположные явления. Пытаются понять преимущества П размножения
(ГФ+РП) перед БП, в то время, как необходимо понять преимущества
РП, перед ГФ (см. табл. 1). Для чего П процесс - понятно: он источник
разнообразия. Необходимо объяснить - что дает ДИФ? И хотя уже поняли,
что, если у П способов нет явных преимуществ перед БП, то должны
быть существенные эволюционные преимущества23,
все равно проблему пола продолжают решать, как проблему размножения,
а не эволюции.
ИДЕЯ АСИНХРОННОЙ (ДИХРОННОЙ) ЭВОЛЮЦИИ
В рамках адаптогенеза Дарвина, эволюция системы следует
за изменением среды и идет методом проб и ошибок. Поэтому выгоднее
пробовать не на целой системе, а только на ее части. Для этого необходимо
разделить систему на две части: одну - основную, более ценную, убрать
подальше от среды, с тем, чтобы лучше сохранить прошлое, а вторую:
“экспериментальную”, менее ценную - выдвинуть поближе к среде, чтобы
“узнать”, что требуется в настоящем и как измениться в будущем.
Такая консервативно-оперативная специализация частей достигается
их последовательной (каскадной), дихронной (для бинарных систем)
эволюцией: все новые признаки появляются сначала в оперативной подсистеме
(у мужского пола), проходят там проверку, после чего - передаются
в консервативную (женскому полу)24.
В 1972 г. я обобщил эту идею для консервативно-оперативной
трактовки целого ряда бинарных эволюционирующих следящих систем
от молекулярного уровня организации до популяционного и социального:
ДНК-белки, А-ПХ, ядро-цитоплазма, женский-мужской пол, подкорка-кора
головного мозга и т.д. Была также высказана гипотеза о том, что
все ДИФ адаптивных систем можно рассматривать, как оперативно-консервативные
специализации, определяющие последовательность попадания информации
от среды в подсистемы25.
На такой базе были созданы изоморфные теории дихронной эволюции
полов (ТДЭП)26,
асимметрии организмов и мозга27,
обладающие исключительным для биологических теорий объяснительным
и предсказательным потенциалом28.
Позже была сделана попытка распространить эту идеологию на А-Г ДИФ
генома29.
Делится ли РП популяция на женский - мужской пол, билатеральный
организм, мозг или др. парные органы на левую - правую половины,
общество на правшей - левшей, геном на А - ПХ, или ПХ на Х- и Y-хр,
все эти ДИФ зиждутся на одном и том же, главном для эволюционирующих
систем, принципе специализации: по сохранению (С) и изменению
(И) системы. Во-первых, наличие С и И - два главных условия
эволюции. Если одно из них отсутствует, то нет эволюции: система
либо исчезает, либо стабильна. Во-вторых, их соотношение, И/С, характеризует
эволюционную пластичность системы. В-третьих, эти условия альтернативны:
чем больше И, тем меньше С, и наоборот, т.к. они дополняют друг
друга до единицы: С + И = 1. Поэтому без специализации
подсистем, системе приходится выбирать некий компромиссный оптимум
И/C, в то время, как при их дополняющей друг друга специализации
возможно увеличить одновременно и то и другое. Скажем, при изогамии
каждая гамета выполняет и консервативную (обеспечение зиготы ресурсами)
и оперативную (поиск партнера) функции. Имея одинаковые средние
(с) размеры, изогаметы и то и другое делают посредственно. Дифференциация
по размеру позволяет мелким (м) лучше осуществлять поиск,
а крупным (к) - обеспечение ресурсами, тогда сочетание к-м
выгоднее чем с-с. В этом эволюционное преимущество ДИФ.
В каждой из приведенных выше пар первая подсистема
консервативная, главная, удаленная и защищенная от среды, вторая
- оперативная “экспериментальная”, теснее связанная со средой. Поэтому
управляющая информация от среды попадает сначала в оперативную подсистему,
оттуда в консервативную. Это приводит к тому, что эволюция любого
признака происходит в них дихронно (асинхронно): у оперативных
она и начинается, и кончается раньше, чем у консервативных.
Следовательно, согласно теории, новые признаки появляются
сначала у мужского пола, потом (спустя много поколений) передаются
женскому30; новые
функции являются ведущими, сначала на правой стороне тела, потом
- на левой; доминирующие центры управления ими возникают сначала
в левом полушарии мозга, потом транслоцируют в правое31.
Точно так же, новые гены должны появляться прежде в половых хромосомах,
потом, в аутосомах32.
ТДЭП. Появление дихронизма
и фазы эволюции признака.
Половая ДИФ - экономная форма информационного контакта
РП популяции со средой, позволяющая, благодаря дихронной эволюциии,
все новые признаки проверять у мужского пола, прежде чем передавать
их женскому (рис. 1, 2).
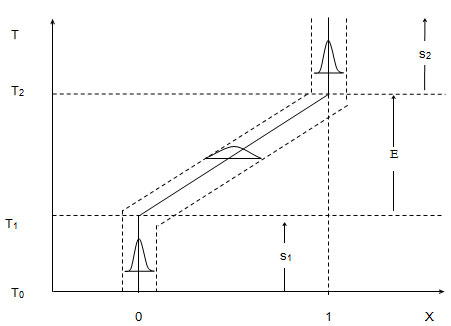
Рисунок 1. Эволюция признака (0–1) у нераздельнополых
форм
По оси абсцисс: `Х - среднепопуляционный генотип по
данному признаку, 0 - до эволюции, 1 - после
эволюции. По оси ординат: Т - время в филогенетическом
масштабе, Т1 - начало, Т2
- конец эволюции признака. Э - стадия эволюции
признака, с1 - доэволюционная,
с2 - послеэволюционная фаза
стабильной стадии признака. Пунктирные линии, параллельные траектории
и маленькие кривые распределения между ними, показывают величину
дисперсии генотипов в популяции в разных фазах.
Мужской и женский пол по разному реагируют на изменения
среды, в частности, появление экологического дифференциала. В 1974
г. мной была выдвинута гипотеза о более широкой норме реакции женского
пола, по сравнению с мужским, позволившая успешно предсказать большую
конкордантность мужских пар монозиготных близнецов и женских пар
- дизиготных33.
Более широкая норма реакции женских особей позволяет им, на
базе старого генотипа, только за счет онтогенетической пластичности
создать более адаптивный фенотип и покинуть зону отбора. Узкая норма
реакции мужских особей такой возможности не дает. Это приводит к
тому, что отбор действует в основном на мужской пол: падает его
численность и меняется распределение генотипов, т.е. начинается
эволюция. Значит одна и та же информация от среды модифицирует женский
пол и элиминирует - мужской. Иными словами, ПД по норме реакции
обеспечивает повышенную фенотипическую пластичность женскому полу
в онтогенезе, и - генотипическую пластичность мужскому полу в филогенезе,
т.е. опережающую эволюцию34.
Значит, женский пол преобразует экологическую информацию во временный
фенотипический ПД, а мужской пол - заплатив за нее своей численностью
- в ГПД, с тем, чтобы после проверки передать её женскому полу “бескровным”
внутренним путем. Стало быть ГПД становится для женского пола движущим
потенциалом, вместо экологического дифференциала, позволяющим ему
новую информацию получать не от среды, а от мужского пола, минуя
отбор. В этом эволюционный смысл половой ДИФ и главное
преимущество РП.
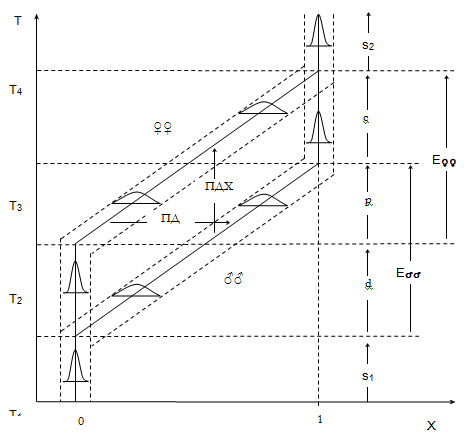
Рисунок 2. Асинхронная эволюция признака (0 – 1)
у мужского (♂♂) и женского (♀♀) пола
Т1 —начало, Т3
—конец эволюции признака у мужского пола. Т2
- начало, Т4 - конец эволюции признака
у женского пола. д - дивергентная, п - параллельная,
к — конвергентная фазы эволюции. Э♂♂
- эволюция признака у мужского пола. Э♀♀
- эволюция признака у женского пола. ПДМ - половой
диморфизм, ПДХ - половой дихронизм. Остальные обозначения
те же, что и на рис. 1.
Эволюция любого признака у РП форм, согласно теории,
включает три фазы.
В первой, дивергентной фазе эволюционирует только мужской
пол, т.к. новая информация от среды попадает только к нему.
Появляется ГПД, который растет из поколения в поколение (фаза роста
ГПД). Длительность дивергентной фазы, величина асинхронии, или П
дихронизм, равна отставанию женского пола, или опережению - мужского
(рис. 2). Эта временна¢я “дистанция” необходима для проверки
новых признаков у мужского пола. Но дивергенция полов не может
продолжаться бесконечно, т.к. иначе наступит репродуктивная изоляция.
Включается механизм оттока информации от мужского пола
к женскому. Начинает изменяться женский пол. Это вторая фаза
эволюции признака - параллельная, когда эволюционируют с одинаковой
скоростью оба пола. Устанавливается стационарный режим для
ГПД, который остается постоянным до конца фазы.
Третья фаза эволюции - конвергентная, в ней эволюционирует
только женский пол. Начинается она, когда на мужской пол перестает
действовать экологический дифференциал, а на женский пол продолжает
действовать ГПД. Происходит уменьшение и утрата (релаксация)
ГПД. Диморфный в процессе эволюции признак становится
снова мономорфным и стабильным. На этом эволюция
признака кончается. Это значит, что РП, которую мы привыкли считать
эффективным способом размножения, скорее - эффективный способ эволюции35.
Поэтому естественно, что новая трактовка коренного понятия - пола
(ДИФ) позволяет, соответственно, по-новому трактовать и все производные
от него понятия: ПД, СП, ПХ, половых гормонов и др., которые также
приобретают эволюционный смысл.
Половой диморфизм
Едва ли не единственное объяснение возникновения ПД
- теория полового отбора Дарвина36.
Но, объяснив общее явление ПД как следствие частного механизма полового
отбора, Дарвин допустил методологическую ошибку. Трактующая теория
всегда должна быть шире трактуемого явления. В этом причина
слабости теории полового отбора. Она не могла объяснить существование
ПД у растений, у которых нет полового отбора, у животных по признакам,
не имеющим отношения к половому отбору, ничего не могла предсказывать37.
Согласно теории, ГПД - следствие дихронной эволюции
полов, поэтому может быть только по эволюционирующим признакам.
Он возникает всегда при эволюции любого признака, как “дистанция”
между полами при любом отборе: естественном, половом, искусственном.
При этом вектор ГПД (от женской формы признака к мужской) показывает
направление эволюции этого признака.
Если новая информация (Iн) уже попала к мужскому
полу (“карантин”), но еще не попала к женскому, или старая информация
(Ic) уже утрачена мужским полом, но еще сохраняется у
женского (“архив”), их сумма и составляет ГПД. Следовательно информация,
содержащаяся в мужском геноме: Iм = Io +
Iн , а в женском: Iж = Io +
Ic , где: Io - общая информация, присутствующая
и там и там (в том числе и о первичных половых признаках обоих полов).
Отметим, что при смешении двух популяций (видов, рас, этносов),
общая информация перемешивается в первом же поколении, а новая
и старая - остаются обособленными на время полового дихронизма.
Такое представление легко объясняет, в частности, различия
межвидовых, межрасовых или межнациональных реципрокных гибридов,
связанные с направлением скрещивания, т.к. у реципрокных гибридов
одинакова только общая часть информации, а Iн и Ic
они получают от разных форм (например, мул и лошак).
Если бы потомки от отца и матери получали одинаковую генетическую
информацию, то не должно было быть никаких реципрокных эффектов.
ТДЭП позволяет делать целый ряд успешных, легко проверяемых,
предсказаний, естественно и просто объясняет с единых позиций
множество, непонятных ранее, явлений и фактов. Например, эволюция
большинства видов позвоночных сопровождалась укрупнением размеров,
а многие виды насекомых и паукообразных, наоборот - мельчали. Стало
быть, согласно теории, самцы должны быть крупнее самок чаще у крупных
позвоночных, а мельче самок - у мелких форм насекомых и паукообразных.
Это так и есть. Та же тенденция наблюдается и в классе млекопитающих,
и в отряде приматов и в других таксонах.
Подтверждает теорию также продвинутость мужского пола,
по сравнению с женским по всем селекционным признакам у культурных
растений и животных. Самцы дают намного больше мяса и лучшего качества,
чем самки, у них выше оплата корма и динамика роста. В тонкорунном
овцеводстве, пушном звероводстве, коневодстве, оленеводстве, шелководстве
и в производстве конопли мужские особи превосходят женских по всем
селекционным качествам.
По признакам же, присущим фенотипу только одного пола,
теорию можно проверить по реципрокным эффектам, поскольку она предсказывает
наличие “отцовского эффекта” (доминирование отцовской породы)
у реципрокных гибридов по всем дивергентно-эволюционирующим, признакам,
в том числе и чисто женским, и материнского эффекта по конвергентно-эволюционирующим38.
Отцовский эффект установлен по инстинкту насиживания, скороспелости,
яйценоскости и живому весу у кур, по динамике роста, числу позвонков
и длине тонкого кишечника у свиней, по удою молока и продукции молочного
жира у крупного рогатого скота39,
по алкоголизму у человека40
и др. Все эти признаки явно новые, ибо являются результатом селекции
или социальных причин. Наличие отцовского эффекта по удою и яйценоскости
означает не что иное, как более высокую генотипическую “удойность”
у быков и “яйценоскость” у петухов, чем у коров и кур, тех же пород.
Представления ТДЭП об обособленности новой и старой
информации на протяжении многих поколений, с учетом исторических
процессов смешения этносов, таких как: переселения (участвуют оба
пола от обоих этносов), завоевания (завоеватели - мужчины, а в покоренном
этносе - оба пола), депортация женщин завоеванного этноса к себе
на родину (2 женских этноса, 1 мужской) и т.д., позволяют объяснить
и другую группу непонятных явлений - в антропологии.
О.М.Павловский в туркменской популяции методом обобщенного
портрета, обнаружил четкий ПД: женские портреты укладывались в один
тип, мужские - только в два типа41.
Аналогичное явление открыл Р.М. Юсупов в краниологии башкир: мономодальное
распределение признаков женских черепов и тетрамодальное - мужских42.
Н.А. Долинова и Л.Г. Кавгазова, соответственно, в удмуртской
и болгарской популяциях, обнаружили ПД по дерматоглифике, когда
у женщин форма одного соседнего этноса, у мужчин - другого43.
Ни одна из существующих теорий, кроме ТДЭП, эти факты не может объяснить.
АУТОСОМНО-ГОНОСОМНАЯ ДИФФЕРЕНЦИАЦИЯ ГЕНОМА
Когда в 1965 г. идея дихронной эволюции впервые была
использована мной для решения проблемы пола, я понял, что она лежит
и в основе аутосомно-гоносомной дифференциации генома, и в заключение
статьи писал: “в хромосомном наборе ПХ выполняют роль оперативной
памяти, а А- постоянной, поэтому ПХ (в первую очередь Y-хр) являются
“воротами” для изменчивости в наследственность”44.
Сейчас это теоретическое предсказание полностью подтверждается серией
экспериментальных исследований скорости замещения нуклеотидных пар45.
Когда система разделена на подсистемы, то прежде всего
встает вопрос о последовательности попадания управляющей информации
от среды. Она всегда попадает сначала в оперативную подсистему,
оттуда передается в консервативную46.
Так, при половой ДИФ: cреда–мужской пол–женский пол. При ДИФ мозга:
среда– левое полушарие– правое полушарие. Если хромосомная ДИФ действительно
изоморфна половой, то этот изоморфизм можно использовать для выявления
каскадной эволюции и для выяснения эволюционной роли А- и ПХ. Для
этого достаточно сопоставить, вскрытые теорией, разные эволюционные
роли мужского и женского пола в получении информации от среды, ее
преобразовании в фено- или генотипическую и передаче потомкам,
т.е их “информационное поведение” с алгоритмами передачи хромосом.
Алгоритмы хромосом
и их сопоставление с фазами эволюции
В информационном поведении хромосом можно выделить алгоритмы:
“вертикальные” - передача самих хромосом из поколения
в поколение, и “горизонтальные” - поступление информации от
среды, передачи между хромосомами и утрата её, в виде генетических
процессов мутагенеза, кроссинговера, транслокаций, переноса эписомами,
вирусами, плазмидами, мобильными генами и т.д. Поведение хромосом
определяют прежде всего три вертикальных алгоритма. 1. Стохастический,
когда хромосомы гомологичной пары попадают сыну или дочери чисто
случайно. Этим алгоритмом передаются А- и Х-хр гомогаметного пола.
2. Ипси-алгоритм, когда хромосома переходит от родителя только к
потомку того же пола. Это алгоритм Y-хр. 3. Контра-алгоритм
- осуществляет переход от родителя к потомку противоположного пола.
Это алгоритм Х-хр гетерогаметного пола.
Стохастический алгоритм имеет дело только с общей для
обоих полов генетической информацией. Перемешивая гены при
каждом оплодотворении, он максимизирует мономодальное генотипическое
разнообразие, гомогенизирует, выравнивает все скачки потенциалов,
поэтому на его основе не может возникнуть ГПД. Он самый древний
алгоритм, который существовал еще до появления дифференциации, реализует
только программы репродукции и рекомбинации.
Нестохастические алгоритмы возникли позже РП и имеют
дело с генетической информацией, разной у мужского и женского пола
- ГПД. Они создают, поддерживают и регулируют ГПД. При этом, ипси-алгоритм
- инициатор программы ДИФ, передавая информацию в пределах одного
пола, создает информационный потенциал между полами - ГПД, увеличивает
или уменьшает его величину. Контра-алгоритм, как и стохастический,
переносит информацию от одного пола другому, поэтому тоже нивелирует
потенциалы, но в отличие от него, не сводит ГПД к нулю, а сохраняет
его постоянство. Сочетание ипси-контра алгоритмов обладает важной
особенностью: оно позволяет создавать и сохранять между подсистемами
определенный ГПД, поддерживать его постоянство и регулировать
его в зависимости от условий среды. При этом, контра-алгоритм
выступает как стабилизатор ГПД (отрицательная обратная связь), а
ипси-алгоритм - как регулятор (положительная обратная связь)47.
Сочетание ипси-контра связей имеет общее, кибернетическое
значение для всех случаев, когда требуется создавать и поддерживать
определенную “дистанцию” между подсистемами. Такая схема управления
лежит в основе и другого, не имеющего до сих пор объяснения, фундаментального
явления нейробиологии - нервного перекреста (управление полушариями
мозга противоположной стороны тела)48.
Такая же схема должна быть и в регуляции П гормонов у мужского
и женского пола.
Итак, в какой последовательности действуют алгоритмы,
для обеспечения дихронной эволюции полов?
1. Чтобы в дивергентной фазе эволюционировал только
мужской пол и при этом возникал и увеличивался ГПД, новая информация
от среды должна поступать только через ипси-алгоритм. Совершенно
очевидно, что задачи этой фазы - доставку новой информации в мужской
геном, ее накопление там в виде ГПД для первичной проверки - может
реализовать только Y-хр.
2. Чтобы в параллельной фазе эволюционировали оба пола,
а ГПД оставался постоянным, необходим отток новой информации из
Y-хр в женский геном. Это может делать только контра-Х-хр.
3. Чтобы в конвергентной фазе эволюционировал только
женский пол и при этом уменьшался и исчезал ГПД, необходимо прекращение
поступления новой информации из среды в Y-хр и продолжение ее оттока
через Х-хр в женский геном.
Эволюционный маршрут
генов по хромосомам
Поскольку для проверки гена в Y-хр требуется много поколений,
то новый ген должен оставаться там необходимый срок, и только после
этого получить возможность покинуть ее. При частичной конъюгации
Х- и Y-хр, как у животных, так и у растений (гуппи, меландриум),
только часть Y-генов находится в конъюгирующем участке49.
Если бы новые гены поступали непосредственно в этот участок, то
они могли бы сразу попасть в женский геном, но по логике ТДЭП, этого
нельзя допустить. Следовательно, “вход” и “выход” Y-хр должны быть
удалены друг от друга. И время, которое ген тратит на перемещение
вдоль Y-хр, чтобы попасть в конъюгирующий с Х-хр участок, равно
П дихронизму. Поэтому только гены, прошедшие “карантин” в Y-хр,
попав в конъюгирующий участок, начинают передаваться неравным
кроссинговером между Y- и Х-хр в женский геном. В Х-хр, видимо,
так же, как и в Y-, вход и выход удалены друг от друга, поэтому
передвигаясь вдоль Х-хр, “молодой” ген проходит вторые испытания
в мужском геноме, будучи в гемизиготном состоянии (рис. 3). Значит,
каждый новый ген, прежде чем попасть в А-, проходит двойную проверку
в ПХ: сначала в дивергентной фазе в Y-хр, потом в параллельной фазе
в контра-Х-хр. Но т.к. рецессивные Х-гены проявляются только у мужского
пола, то отбору подвергается только он.
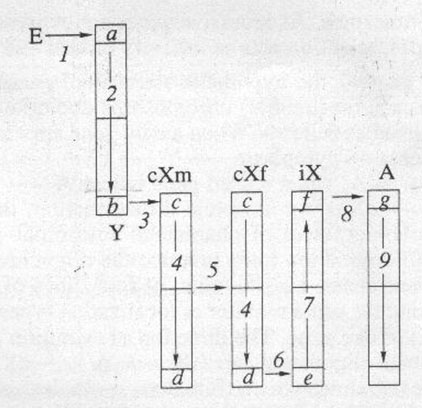
Рисунок 3. Гипотетическая схема маршрута нового гена
по половым хромосомам в дивергентной фазе эволюции признака
С - среда (цитоплазма). Участки хромосом:
а - вход Y-хромосомы, не участвующий в кроссинговере с
cХм-хромосомой; b - выход Y-хромосомы, участвующий в неравном
кроссинговере с cХм-хромосомой; c - вход cХм-хромосомы,
участвующий в неравном кроссинговере с Y-хромосомой; d - выход
cХм-хромосомы, участвующий в неравном кроссинговере с кХf-хромосомой;
f - вход iХ-хромосомы, участвующий в неравном кроссинговере
с cХf-хромосомой; е — выход iХ-хромосомы, транслокации
генов на аутосомы; g - вход аутосом, приема транслокаций.
Переходы генов: 1 - из cреды в Y-хромосому
(мутагенез); 2 - по длине Y-хромосомы из неконъюгирующего
(a) участка в конъюгирующий (b); 3 - неравный
кроссинговер Y–cХм; 4 - по длине cХ-хромосомы в мужском
и женском геноме; 5 - вертикальный cХ-алгоритм (отец –
дочь); 6 - неравный кроссинговер: cХм–iХ; 7
- по длине iХ-хромосомы; 8 - транслокация (плазмиды, вирусы)
iХ-хромосома – аутосомы. 9 - по длине аутосом.
Итак, можно попытаться нарисовать общую дедуктивно-гипотетическую
картину переходов генов при каскадной (последовательной) эволюции
хромосом генома. При появлении нового гена: среда – цитоплазма
– Y – cХм – cХж – (iХ?) – А, а при утрате отработавшего
гена: А – iХ – cХж – cХм. Стало быть, точно так, как
при П ДИФ существует дихрономорфизм признаков, так и при А-Г
ДИФ должен существовать олигохрономорфизм генов, т.е. три-четыре
разных времени появления и столько же форм локализации (Y,
cХ, А, иХ,) одного и того же гена. При этом векторы эволюции
хромосомного олигоморфизма (iХ – А – cХ – Y), как и ПД (ж – м),
всегда направлены противоположно потоку информации (Y – cХ–
А – iХ, м – ж) и могут служить “компасом” эволюции (рис. 2). Зная
маршрут генов, можем вскрыть принцип их локализации по хромосомам.
Какие гены локализованы
в аутосомах, какие - в половых хромосомах?
На этот вопрос классическая хромосомная теория ответа
не дает. Но, по идее и названиям хромосом (половые, гоносомы) подразумевается,
что в ПХ должны быть гены признаков, связанных с полом и размножением,
а в А- - неполовых, соматических, т.е. критерий локализации - репродуктивный.
Новая же концепция дает четкий ответ: в ПХ должны быть гены эволюционирующих
признаков, а в аутосомах - константных (стабильных), т.е критерий
- эволюционный (табл. 2). Каковы факты?
Совершенно очевидно, что все 30 Y-генов окраски гуппи
(кроме одного А-гена)50,
а также Y-гены волосатости ушей и перепонок между пальцами ног человека,
эволюционно-новые соматические мутации, не имеющие прямого отношения
к репродуктивной функции. В то же время гены “самых репродуктивных”
признаков - первично половых (гамет, гонад, гениталий) локализованы
в аутосомах. Стало быть эти факты противоречат существующей теории
и подтверждают новую концепцию, т.е. “водораздел” между А- и Г-генами
проходит не по размножению, а по эволюции.
Таблица 2. Какие гены локализованы в аутосомах - какие
в половых хромосомах?
|
|
Комбинации признаков*
|
|
Локализация генов
|
С,К
|
Р,К
|
С,Э
|
Р,Э
|
|
По классической теории
|
А
|
ПХ
|
А
|
ПХ
|
|
По новой концепции
|
А
|
А
|
ПХ
|
ПХ
|
|
Половой диморфизм
|
-
|
РПД
|
ЭПД
|
ПД+ЭПД
|
|
Основа пол. диморфизма
|
-
|
Фенотипич.
|
Генотипич.
|
Фен+ген
|
* С - соматические, К - константные, Р
- репродуктивные, Э - эволюционирующие.
Далее, дихронная эволюция означает, что в генофонде
РП популяции, по эволюционному “возрасту”, локализации по хромосомам
того и другого пола, могут быть три группы генов.
1. Сугубо мужские Y- и Х-гены (“футуристических” признаков
- новые, молодые, “завтрашние”), которые появились у мужского
пола, но еще не прошли проверку, не попали в А- и не стали общим
достоянием.
2. Общие гены (рабочие, актуальные, “сегодняшние”),
это основная масса генома - А-гены, присутствующие одновременно
у того и другого пола.
3. Сугубо женские Х-гены (старые, отработавшие уже в
А-, “вчерашние”), которые уже утрачены мужским полом, но ещё сохраняются
у женского, в виде атавистических признаков в ожидании элиминации.
Необходимость существования последних вытекает, во-первых, из теории,
во-вторых, следует из целого ряда известных явлений и фактов, которые
иначе трудно объяснить51.
Локализованы эти гены, видимо, в специальном участке Х-хр (а может
и А-) и в мужской геном попадают только для элиминации.
Так как Г-гены эволюционируют, а А-гены - стабильны,
то соотношение Г/А отражает эволюционную пластичность генома:
в популяции, долго живущей в стабильной среде, нет Г-генов (Г/А=0)
и чем изменчивее среда, тем больше Г/А. Значит существует,
зависящее от среды равновесие [A] <=> [Г] (аналог
СП: [Ж] <=> [М]), которое в оптимальной среде сдвигается
влево, в экстремальной - вправо. Отметим, что при смешении популяций
А-гены перемешиваются у потомков первого же поколения, а Г-гены,
остаются обособленными на протяжении П дихронизма (см. рис.4.).
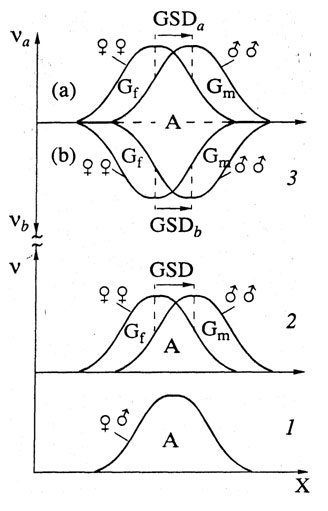
Рисунок 4. Аутосомные (А) и гоносомные (Г) гены
По оси абсцисс: генотипы (G) по данному признаку; по
оси ординат: их частоты (n) в популяции. 1. В популяции,
долго живущей в стабильной среде, все гены локализованы в А, поэтому
кривые распределения мужских и женских генотипов совпадают (Г/А=0,
ГПД=0, ФПД=0, “завтрашние” и “вчерашние” гены отсутствуют, признак
стабилен и не бывает реципрокных эффектов). 2. В популяции,
живущей в изменчивой среде, помимо А-генов, есть и Г-гены: мужские
(Гм) и женские (Гж), поэтому
эволюционная пластичность, Г/А=0, ГПД=0, признак эволюционирует
и могут быть реципрокные эффекты). 3. При смешении популяций
а и б, барьер между А-генами (пунктирная линия)
исчезает, т.е. у гибридов первого же поколения А-гены перемешиваются.
Барьеры же между Г-генами (жирные линии), остаются на протяжении
П-дихронизма. Поэтому могут существовать по два типа ГПД
и реципрокных гибридов обоих полов (мул, лошак).
Следовательно и ПД, в зависимости от хромосомной и половой
принадлежности его генной основы, может быть трех видов. 1. При
появлении любого признака, сугубо мужские гены образуют, обусловленный
дихронизмом (т.е. гоносомный) временный эволюционный ГПД “футуристической”
природы. Как только эти гены, достигнув аутосом, становятся общими,
ГПД по ним исчезает. 2. Но, по признакам, имеющим разное селективное
значение для мужского и женского пола, ГПД подхватывается
половыми гормонами и превращается в аутосомный постоянный ФПД. Значит
по таким признакам (первично- и вторично-половым), ГПД отсутствует,
а наличие и степень выраженности ФПД определяет гормональный пол
организма. 3. При утрате любого признака, сугубо женские гены также
образуют, обусловленный дихронизмом, временный эволюционный ГПД,
но “атавистической” природы. Он возникает, когда общий
ген, становится сугубо женским, а исчезает при элиминации гена в
контра-Х-хр. Постоянный ФПД, является конститутивным (базовым),
тогда как эволюционные ГПД - факультативны, могут возникнуть только
при наличии конститутивного. Если же эволюционирует репродуктивный
признак, то по нему может быть двойной ПД - и постоянный и эволюционный.
Как уже говорилось, при изменении среды, из-за разной
нормы реакции, женский пол покидает зону отбора, а мужской - остается
в ней. Стало быть необходимо, чтобы у них заранее (в стабильной
фазе) существовал ГПД по норме реакции. Причем, генетическая информация
о широкой норме реакции должна передаваться только по женской линии,
а об узкой—только по мужской. Это также могут делать только ипси-алгоритмы,
т.е. Y-хр и ипси-участок Х-хр.
Но почему именно у мужского пола узкая норма реакции
и оперативная специализация, а не у женского и бывает ли наоборот?
Реверсия полового диморфизма
при полиандрии
Полиандрия, при которой самка спаривается с несколькими
самцами, т.е. имеет более широкое “сечение канала” передачи генетической
информации потомству, чем самец, встречается у беспозвоночных, рыб,
птиц, млекопитающих. При этом часто наблюдается реверсия ПД (самки
крупнее самцов, ярче окрашены, самцы строят гнездо, насиживают яйца
и заботятся о выводке, отсутствует борьба за самку). При полигинии
- картина обратная. Это значит, что направление дихронии, соотношение
скоростей эволюции, зависит от направления полигамии, т.е. соотношения
“сечений каналов” связи. В строго моногамной популяции самцы и самки
имеют одинаковое “сечение канала”, т.е. число отцов и матерей
равно, тогда одинаковы также и дисперсии ипси-хромосом у сыновей
и у дочерей. В случае полигинии, когда отцов меньше, чем матерей,
дисперсия Y-хр у сыновей меньше, чем ипси-Х-хр у дочерей. В случае
полиандрии - все наоборот. Значит, дисперсия Y-хр у сыновей пропорциональна
числу отцов, а дисперсия ипси-Х-хр у дочерей - числу матерей. Кроме
того, Y-хр запускает синтез тестостерона, концентрация которого
определяет ПД. С другой стороны, как было показано выше, соотношение
норм реакций мужского и женского пола определяет направление ГПД
по любому признаку.
Следовательно, можно высказать гипотезу, о том,
что норма реакции обратно пропорциональна концентрации тестостерона
в организме, и, построив причинно-следственную зависимость, связать
направление ГПД, т.е. соотношение скоростей эволюции мужского и
женского пола (Эм/Эж) c соотношением
их “сечений каналов” (Сж/Cм). Сж/Cм
~ Nотцов/Nматерей ~ sY/sиX
~ Тм/Tж ~ Нж/Нм
~ Эм/Э ж , где: N - численность, sY,
sиX - дисперсии Y-хр у сыновей и ипси-Х-хр
у дочерей, Тм , Тж - концентрации тестостерона,
Нм, Нж - нормы реакций, ~
- знак пропорциональности. Значит, роль эволюционного “авангарда”
всегда получает более полигамный пол, а “арьергарда” - моногамный.
А почему полигиния широко распространена в природе, а полиандрия
- экзотика, объясняется потенциально большими репродуктивными
возможностьями мужского пола (в конечном счете большим числом
гамет). Ведь строго говоря, полиандрии, как таковой, не может быть,
в лучшем случае это только олигоандрия: ибо возможности женского
пола в этом смысле ограничены. Такая трактовка позволяет делать
четкое предсказание: у отцовских полусибсов (потомков одного отца
и разных матерей) жизнеспособность должна быть выше у дочерей, а
у материнских полусибсов, наоборот, - у сыновей.
Эволюционная логика
аутосом и половых хромосом
Аутосомы, будучи консервативной памятью РП генома (аналог
женского пола в популяции) и хранилищем стабильных, общих для обоих
полов, генов, нацелены на его сохранение. Эволюционно - это самые
старые хромосомы, содержащие фундаментальную видовую информацию.
Они выполняют более древние программы РЕП и РЕК. Передаваясь стохастически,
они перемешиваются в каждом поколении, обеспечивая максимальное
разнообразие генотипов, т.е. наилучшим образом реализуют программы
П процесса, по которым наивысшие достижения у ГФ. В этом смысле
они скорее “рекомбинационные”.
Половые хромосомы, являясь оперативной памятью или экспериментальной
подсистемой генома, нацелены на его изменение (аналог мужского пола
в популяции). Точно так, как новый признак не появляется в
женском фенотипе, не пройдя проверку в мужском, так и новый ген
не появляется в А-, не пройдя проверку в ПХ. Главное назначение
ПХ - создание дихрономорфизма для эффективной эволюции. В филогенезе
ПХ появились позже РП, т.е. они значительно “моложе” А-. Запуская
и выполняя программу ДИФ, они формируют в популяции консервативно-оперативные
подсистемы и распределяют между ними роли в зависимости от полигинии
- полиандрии. Для этого снабжают более моногамный пол широкой нормой
реакции, а более полигамный - узкой независимо от типа гаметности52.
Тем самым создают две обособленные и дихронно эволюционирующие подсистемы
с информационными барьерами между ними. Регулируя скорости горизонтальных
переходов (вдоль хромосом и между ними), регламентируют и дозируют
попадание новой информации в женский геном. В них находятся, главным
образом, эволюционирующие гены, как приобретаемые, так и утрачиваемые.
Их деятельность, будучи направлена против программы РЕК (запрет
комбинаций мм, жж), ухудшает вдвое разнообразие генотипов, возможное
в результате П процесса и в этом смысле они скорее “антиполовые”,
чем “половые”. Поэтому логичнее было бы, с учетом
их роли, назвать их - “эволюционными”.
Y-хромосома - связующее звено между геномом и средой
(цитоплазмой). Кстати, на дрозофиле было показано, что критерием
наступления репродуктивной изоляции между разными расами является
несовместимость Y-хр одной расы с цитоплазмой - другой53.
Y-хр - “ворота” для новой информации в геном. Она трансформирует
экологическую информацию в генетическую, т.е. создает новые гены,
мутации. Y-хр - запускает мужские П гормоны и через них определяет
норму реакции мужского пола; содержит “завтрашние” гены; инициатор,
акселератор и регулятор П дихрономорфизма; первый “испытательный
полигон” и “карантинный изолятор” для новых генов. Поэтому больше
основния ее назвать “экологической”.
Контра-Х-хромосома - переносчик генов, связующее звено
между Y-хр и женским геномом - “транспортная”; стабилизатор, релаксатор
и ликвидатор П дихрономорфизма в филогенезе; второй полигон, где
молодые гены, будучи в гемизиготном состоянии у мужского пола, проходят
испытания в онтогенезе. По этой же причине (подверженности интенсивному
отбору), она является, возможно, местом элиминации ставших бесполезными,
или вредными, генов, поступающих из А-.
Ипси-Х-хромосома (специальный ее участок) - должна быть
прежде всего, связана с определением П гормонов и нормы реакции
женского пола (в зависимости от типа полигамии) и хранением сугубо
женских генов. В Х-хр должна быть больше доля модификационных генов
количественных признаков.
Эволюция определения
пола и ПД. Гормональный и психический пол
В процессе прогрессивной эволюции определение пола закономерно
переходит от генного (у ГФ) к хромосомному (у РП) и геномному (у
пчел). Параллельно повышается уровень ДИФ и происходит “экспансия”
ПД: у БП форм он отсутствует, у ГФ существует ПД только на уровне
первичных половых признаков (гамет, гонад, гениталий), у РП моногамов
появляется организменный ПД (вторичных половых признаков), у РП
полигамов - популяционный, включающий ПД по численности (СП) и дисперсии
полов, а у пчел - появляются ПД генома (гапло- диплоидия) и новая
консервативно-оперативная дифференциация на касты, при которой существуют
две экологические подсистемы. Трутни других (богатых) семей, приносящие
генетическую информацию (от ближней среды), и рабочие пчелы - от
медоносов (дальней). Стало быть с точки зрения новой концепции,
рабочие пчелы, будучи генетическими самками, выполняют функции второго
экологического (мужского) пола, а способ размножения пчел выступает,
как эволюционно более продвинутая форма. У рыб же с карликовыми
самцами (а также, видимо, у муравьев и термитов с кастами) вторая
консервативно-оперативная ДИФ произошла на базе мужского генома,
но также для доставки экологической информации в женский геном из
двух разных сред.
Такая идеология позволяет выделить экологическую подсистему
в онтогенезе и на других уровнях реализации пола: генный – гормональный
– психический. Тогда возникает новая трактовка андрогенов (а), как
экологических гормонов, “приближающих” систему к среде, а их антагонистов
эстрогенов (э) - “удаляющих”, защищающих от среды. А на поведенческом,
психическом уровне экологической подсистемой, аналогом мужского
пола, выступают левши (л), а женского пола - правши (п). Значит,
по аналогии с СП (м/ж), соотношения а/э и л/п (% левшей) - регуляторы
“дистанции” от среды, а также регуляторы эволюционной пластичности,
которая в оптимальной среде - должна падать, а в экстремальной -
расти54 (см. также
рис.4).
Экспериментальное подтверждение, выдвинутой в 1965 г.
центральной идеи ТДЭП, было получено впервые в 1987 г.55.
Поскольку число клеточных делений в сперматогенезе намного больше
числа делений в овогенезе и ошибки при репликации и репарации ДНК
являются главным источником мутаций для молекулярной эволюции, был
сделан вывод, что все это может привести к большей частоте
мутаций в ПХ по сравнению с А- и было высказано предположение, что
самцы служат генератором мутаций, по крайней мере, в эволюции млекопитающих.
(Тот факт, что уровень, как спонтанного так и индуцированного мутагенеза,
у гетерогаметных, и у гомогаметных самцов выше чем у самок для дрозофилы,
шелкопряда, млекопитающих, в том числе и человека был установлен
давно и неоднократно56).
Сравнительным исследованием замещений нуклеотидной последовательности
А-, Х- и Y-генов человека и мыши (или крысы) было показано, что
самцы являются главным источником мутаций для молекулярной эволюции.
Кроме того, было установлено, что соотношение скоростей эволюции
генов - Y: А : Х = 2,2 : 1 : 0,6 (хорошо согласуется с теоретическим
ожиданием - 2 : 1 : 2/3)57.
В другой работе аналогичной методикой сравнивали Y/X
отношения скоростей замещения нуклеотидных последовательностей в
синонимных генах человека, орангутана, бабуина и беличьей
обезьяны. Было показано, что Y-гены этих видов дивергируют быстрее
и дальше друг от друга, чем Х-гены, т.е., и у высших приматов также
самцы предваряют молекулярную эволюцию58.
Названия обеих работ начинаются словами: “Ведомая самцами эволюция...”.
Таких работ уже много. Я привел только две: первую и вторую (на
мой взгляд, красивую). Эти эксперименты на молекулярном уровне полностью
подтверждают ТДЭП. А поскольку новая концепция ПХ не что иное, как
изоморфная трансформация той же самой идеологии с популяционного
уровня на хромосомный, то с полным основанием можно считать, что
они являются убедительным подтверждением также и основных положений
концепции ПХ.
ЧТО МОЖЕТ ОБЪЯСНИТЬ И ПРЕДСКАЗАТЬ НОВАЯ КОНЦЕПЦИЯ?
Новая концепция пересматривает некоторые, уже вековые,
представления о ПХ и в корне меняет их понимание, порой на противоположное.
Например, их основная роль не репродуктивная - определение пола,
как считали, а эволюционная - создание дихрономорфизма для экономной
эволюции (даже в ущерб размножению!), не обеспечение СП=1:1
и его постоянства, а наоборот, его изменение (регуляция). Становится
понятно явление неравного Y–X кроссинговера, тесная связь Y-хр с
ретровирусами. Изучали участки ДНК Х- и Y-хр мышей методом гибридизации
с ДНК ретровирусов. В составе Y-хр 10 линий мышей, весьма отличающихся
друг от друга, обнаружили ДНК мышиных ретровирусов. В Х-хр их не
было59. Сюда
же относится способность сперматозоидов многих видов связывать
чужеродную ДНК и переносить её при оплодотворении в яйцеклетку.
Явление впервые было описано в 1971 г. по внедрению ДНК вируса SV40
в сперматозоиды кролика60.
Однако открытие не привлекло должного внимания. Позже, оно было
подтверждено и распространено на других животных (мышь, морского
ежа, пчелу, курицу, свинью, корову, человека) с вирусными частицами
и вирусной ДНК. Широкий резонанс вызвали работы, использующие сперматозоиды
в качестве вектора для получения трансгенных животных61.
Это позволяет делать на основе концепции ПХ важный вывод-предсказание:
чужеродную ДНК (или вирусы) связывают не все сперматозоиды, а только
Y-несущие, поэтому среди трансгенных животных должно быть больше
самцов.
Другие, интенсивно исследуемые в последнее время направления,
имеющие прямое отношение к концепции ПХ - “вспышки” мутаций, инсерционный
мутагенез, “прыгающие” гены. Она позволяет объяснить куда и зачем
“прыгают” гены, почему транспозиции мобильных элементов зависят
от экологического стресса (температуры, гамма излучения, химических
веществ, гибридного дисгенеза и др.), почему мутируют одновременно
разные гены, где искать источник мутагенеза - в овогенезе или сперматогенезе,
в А- или ПХ, в Х- или Y-хр и т.д.? Сколько мне известно, эти вопросы
даже не ставятся.
Концепция позволяет по-другому интерпретировать многие
непонятные явления, перечисленные в начале статьи. Скажем, конденсацию
Х-хр в женском геноме (тельца Барра) независимо от типа гаметности,
нужно трактовать не как компенсацию дозы генов (как было уже сказано,
это приводит к алогизму), а как преграду против распространения
у самок новой, еще не проверенной информации. Эухроматиновый характер
Y-хр растений и разбросанные по всем хромосомам нуклеотидные повторы
объясняются относительно поздней эволюцией П ДИФ у растений по сравнению
с животными62. Если
нуклеотидные повторы - для образования новых генов (а до появления
ПХ гены “рождались” во всех хромосомах), то разброс повторов по
хромосомам у растений отражает более раннюю стадию развития у них
П ДИФ.
Далее, Y-хр, как экологическая, должна быть тесно связана
со стрессом. Это может объяснить ряд известных фактов, таких как:
относительно более крупные размеры Y-хр у некоторых этнических (например,
евреев в Германии) или социальных групп (заключенных) у человека,
или ее повышенную дисперсию в зонах высокой сейсмической активности
у грызунов. (Только в последнем случае - это объясняется не высоким
уровнем радиации, или концентрации радона, как считают авторы, а
просто стрессом от частых землетрясений.) В этой связи можно
предсказать изменения размеров и/или дисперсии Y-хр в регионах сильных
и частых землетрясений, других природных бедствий и социальных сдвигов:
геноцида, длительных войн, переселений, голода и пр. А также у интенсивно
эволюционирующих, в результате селекции, животных и растений и т.д.
Настоящая статья - теоретическая63.
Она, главным образом, об эволюционных закономерностях, а не о конкретных
механизмах реализации “горизонтальных” потоков генов - требующих
специального исследования. Здесь же уместно лишь высказать гипотезу
о том, что должна существовать связь между митохондриальной ДНК
и Y-хр, являющейся “воротами” (источником, генератором) не спонтанных
и случайных мутаций, а индуцированных и направленных экологическим
дифференциалом. А это уже программа на будущее.
И в заключение статьи выражаю признательность рецензентам
Ю.Г. Рычкову и Ю.Ф. Богданову за высокую оценку, обсуждение, конструктивную
критику, полезные замечания и советы.
1
Институт проблем экологии и эволюции им. А.Н.Северцова РАН.
2
Эйнштейн А. Физика и реальность // М. 1965. С. 109 (350).
3
Франкфурт У.И., Френк А.М. Джозайя Виллард Гиббс // М., 1964. С.76
(280).
4
Геодакян В.А. Роль полов в передаче и преобразовании генетической
информации // Пробл. передачи информации. 1965. Т. 1, № 1, с. 105-113;
Геодакян В.А. Эволюционная теория пола // Природа. 1991. № 8, с.
60-69; Геодакян В.А. Теория дифференциации полов в проблемах человека
// В кн. Человек в системе наук. 1989. М.: Наука, с. 171-189; Geodakian
V.A. Natural Selection and Sex Differentiation // In: Natural Selection.
Proc. Intern. Symp. Liblice, 1978. (Ed. V.J.A. Novak) Praha, 1978.
P.65-78; Geodakian V.A. Sexual Dimorphism //In: Evolution and Morphogenesis.
Proc. Intern. Symp. Plzen. 1984. (Ed. J. Mlikovsky) Praha-Acad.1985.
P.467-477.
5
Сокращения, встречающиеся в тексте: м - мужской; ж -
женский; П –половой; ПХ - половые хромосомы; ПД
- половой диморфизм: ГПД - генотипический, ФПД
- фенотипический; СП - соотношение полов; хр.
- хромосома; А - аутосома; Г - гоносома; БП
- бесполый; ГФ - гермафродитный; РП - раздельнополый;
программы: РЕП - репродуктивная, РЕК - рекомбинационная;
ДИФ - дифференциации; ТДЭП - теория дихронной эволюции
полов; а - андрогены; э - эстрогены.
6
Геодакян В.А. Половые хромосомы: для чего они? (Новая концепция)
// Докл. Акад. Наук. 1996.Т.346, № 4, с.565-569.
7
Mc. Clung C.E. Notes on the Accessory Chromosome // Anat. Anz. 1901.
Bd. 20. S. 220-226.
8
Vorontsov N.N. The Evolution of the Sex Chromosomes // In: Cyto-
taxonomy and Vertebrate Evolution (Ed. A.B. Chiarelli). 1973, p.
619-657.
9
Grant S., Houben A., Vyskot B., Siroky J., Wei-Hua P., Macas J.,
Genetics of Sex Determination in Flowering Plants // Dev. Genet.
1994. V.15, p. 214-230.
10
Winge O. The Location of Eighteen Genes in Lebistes Reticulatus
// J. Genetics. 1927. V. 18, p. 1-43; Кирпичников В.С. Аутосомные
гены у Lebistes Reticulatus и проблема возникновения генетического
определения пола // Биологический журнал. 1935. Т. IV, №2, с. 343-354.
11
Rice W.R. Degeneration of a Nonrecombining Chromosome // Science.
14 Jenuary 1994. V. 263, p. 230-232.
12
Воронцов Н.Н., Ляпунова Е.А., Иваницкая Е.Ю. Изменчивость половых
хромосом млекопитающих // Генетика. 1978. Т. 15, с. 1432-1446.
13
Phillips S., Birkenmeier E., Callahan R., Eicher E. Male and Female
Mouse DNAs can be Discriminated using Retroviral Probes
// Nature.1982. V. 297, № 5863, p. 241-243.
14
Miyata T., Hayashida H., Kuma K., Mitsuyasu K., Yasunaga T. Male-driven
Molecular Evolution: A Model and Nucleotide Sequence Analysis
// Cold Spring Harbor Symposia on Quantitative Biology. 1987. V.
LII, p. 863-867; 18. Shimmin L.S., Chang B. H-J, Li W-H. Male-driven
Evolution of DNA Sequences // Nature. 22 April 1993. V. 362, p.
745-747.
15
Williams G.C. Sex and Evolution // 1975. Princeton, New Jersey:
Princeton University Press; Maynard Smith J. The Evolution of Sex
// 1978. Cambridge: Camb. Univ. Press; Bell G. The Masterpiece of
Nature // 1982. Berkeley: Univ. of Calif. Press; Bull J.J. Evolution
of Sex Determining Mechanisms // 1983. Menlo Park, California: WA
Benjamin/Cummings; Karlin S., Lessard S. Sex Ratio Evolution //
1986. Princeton, New Jersey: Princeton Univ. Press; Hoekstra R.F.
The Evolution of Sexes. In Stearns S.C. (ed) The Evolution of Sex
and its Concequences // 1987. Basel: Birkhauser Verlag; Michod R.E.,
Levin B.R. (eds) The Evolution of Sex // 1988. Sunderland, Massachusetts:
Sinauer Associates; Dawley R.M., Bogart J.P. Evolution and Ecology
of Unisexual Vertebraes // 1989. Albany: New York State Museum;
Harvey P.H., Partridg L., Southwood T.R.E. (eds). The Evolution
of Reproductive Strategies // 1991. London: The Royal Society; Mooney
S.M. The Evolution of Sex: A Historical and Philosophical Analysis
// 1992. Ph. D. thesis. Boston: Boston Univ.
16
Williams G.C. Sex and Evolution // 1975. Princeton, New Jersey:
Princeton University Press
17
Maynard Smith J. The Evolution of Sex // 1978. Cambridge: Camb.
Univ. Press
18
Bell G. The Masterpiece of Nature // 1982. Berkeley: Univ. of Calif.
Press.
19
Crews D. Animal Sexuality // Scientific American. Jan. 1994, p.
108-114
20
Developmental Genetics. 1994. V. 15, p. 201-312; J. Hered. 1993.
V. 84, p. 321-440.
21
Crow J.F. Advantages of Sexual Reproduction // Developmental Genetics.
1994. V. 15, p. 205-213.
22
Геодакян В.А. Роль полов в передаче и преобразовании генетической
информации // Пробл. передачи информации. 1965. Т. 1, № 1, с. 105-113.
23
Crow J.F. Advantages of Sexual Reproduction // Developmental Genetics.
1994. V. 15, p. 205-213.
24
Геодакян В.А. Роль полов в передаче и преобразовании генетической
информации // Пробл. передачи информации. 1965. Т. 1, № 1, с. 105-113.
25
Геодакян В.А. О структуре эволюционирующих систем // В сб. Проблемы
кибернетики. 1972. М.: Наука, вып. 25, с. 81-91.
26
Геодакян В.А. Эволюционная теория пола // Природа. 1991. № 8,
с. 60-69; Геодакян В.А. Теория дифференциации полов в проблемах
человека // В кн. Человек в системе наук. 1989. М.: Наука, с. 171-189.
27
Геодакян В.А. Асинхронная асимметрия // Журнал высшей нервной деятельности.
1993. Т. 43, вып. 3, с. 543-561.
28
См., например: Симонов П.В., Русалова М.Н., Преображенская Л.А.,
Ванециан Г.Л. Фактор новизны и асимметрия деятельности мозга //
Журнал высшей нервной деятельности. 1995. Т. 45, вып. 1, с. 13-17.
29
Геодакян В.А. Половые хромосомы: для чего они? (Новая концепция)
// Докл. Акад. Наук. 1996.Т.346, № 4, с.565-569.
30
Геодакян В.А. Эволюционная теория пола // Природа. 1991. № 8,
с. 60-69; Геодакян В.А. Теория дифференциации полов в проблемах
человека // В кн. Человек в системе наук. 1989. М.: Наука, с. 171-189.
31
Геодакян В.А. Асинхронная асимметрия // Журнал высшей нервной деятельности.
1993. Т. 43, вып. 3, с. 543-561.
32
Геодакян В.А. Половые хромосомы: для чего они? (Новая концепция)
// Докл. Акад. Наук. 1996.Т.346, № 4, с.565-569.
33
Геодакян В.А. Дифференциальная смертность и норма реакции
мужского и женского пола. Онтогенетическая и филогенетическая пластичность
// Журн. общ. биол. 1974. Т. 35, № 3, с. 376-385.
34
Геодакян В.А. Роль полов в передаче и преобразовании генетической
информации // Пробл. передачи информации. 1965. Т. 1, № 1, с. 105-113;
Геодакян В.А. Эволюционная теория пола // Природа. 1991. № 8,
с. 60-69; Геодакян В.А. Дифференциальная смертность и норма
реакции мужского и женского пола. Онтогенетическая и филогенетическая
пластичность // Журн. общ. биол. 1974. Т. 35, № 3, с. 376-385.
35
Геодакян В.А. Эволюционная теория пола // Природа. 1991. № 8, с.
60-69; Геодакян В.А. Теория дифференциации полов в проблемах человека
// В кн. Человек в системе наук. 1989. М.: Наука, с. 171-189.
36
Дарвин Ч. Происхождение человека и половой отбор // 1953. М.
37
Geodakian V.A. Sexual Dimorphism //In: Evolution and Morphogenesis.
Proc. Intern. Symp. Plzen. 1984. (Ed. J. Mlikovsky) Praha-Acad.1985.
P.467-477; Геодакян В.А. Половой диморфизм // Биол. журн. Армении.1986.
Т.39, № 10, с. 823-834.
38
Геодакян В.А. Эволюционная теория пола // Природа. 1991. № 8,
с. 60-69; Геодакян В.А. Теория дифференциации полов в проблемах
человека // В кн. Человек в системе наук. 1989. М.: Наука, с. 171-189;
Geodakian V.A. Sexual Dimorphism //In: Evolution and Morphogenesis.
Proc. Intern. Symp. Plzen. 1984. (Ed. J. Mlikovsky) Praha-Acad.1985.
P.467-477; Геодакян В.А. Половой диморфизм // Биол. журн. Армении.1986.
Т.39, № 10, с. 823-834; Геодакян В.А. Половой диморфизм и “отцовский
эффект” // Журнал общей биологии. 1981. Т. 42, № 5, с. 657-668.
39
Геодакян В.А. Половой диморфизм и “отцовский эффект” // Журнал общей
биологии. 1981. Т. 42, № 5, с. 657-668
40
Эрман Л., Парсонс П. Генетика поведения и эволюция // 1984. М. “Мир”.
41
Павловский О.М. О чем рассказывает обобщенный фотопортрет? // Наука
и жизнь. 1980. № 1, с. 84-90.
42
Юсупов Р.М. О половом диморфизме и значении женских выборок черепов
в антропологии // В сб. Источники по истории и культуре Башкирии.
1986. Уфа. С. 51-56.
43
Долинова Н.А. Дерматоглифика удмуртов // В сб. Новые исследования
по этногенезу удмуртов. 1989. Ижевск. Уро АН СССР. С. 108-122.
44
Геодакян В.А. Роль полов в передаче и преобразовании генетической
информации // Пробл. передачи информации. 1965. Т. 1, № 1, с. 105-113.
45
Miyata T., Hayashida H., Kuma K., Mitsuyasu K., Yasunaga T. Male-driven
Molecular Evolution: A Model and Nucleotide Sequence Analysis
// Cold Spring Harbor Symposia on Quantitative Biology. 1987. V.
LII, p. 863-867; Shimmin L.S., Chang B. H-J, Li W-H. Male-driven
Evolution of DNA Sequences // Nature. 22 April 1993. V. 362, p.
745-747.
46
Геодакян В.А. О структуре эволюционирующих систем // В сб. Проблемы
кибернетики. 1972. М.: Наука, вып. 25, с. 81-91.
47
Geodakian V.A. Feedback Control of Sexual Dimorphism and Dispersion
// In: Towards a New Synthesis in Evolutionary
Biology. 1987. Proc. Intern. Symp. Praha. Czech. Acad. Sci.
P.171-173.
48
Геодакян В.А. Асинхронная асимметрия // Журнал высшей нервной деятельности.
1993. Т. 43, вып. 3, с. 543-561.
49
Grant S., Houben A., Vyskot B., Siroky J., Wei-Hua P., Macas J.,
Genetics of Sex Determination in Flowering Plants // Dev. Genet.
1994. V.15, p. 214-230; Winge O. The Location of Eighteen Genes
in Lebistes Reticulatus // J. Genetics. 1927. V. 18, p. 1-43;
Кирпичников В.С. Аутосомные гены у Lebistes Reticulatus и
проблема возникновения генетического определения пола // Биологический
журнал. 1935. Т. IV, №2, с. 343-354.
50
Winge O. The Location of Eighteen Genes in Lebistes Reticulatus
// J. Genetics. 1927. V. 18, p. 1-43; Кирпичников В.С. Аутосомные
гены у Lebistes Reticulatus и проблема возникновения генетического
определения пола // Биологический журнал. 1935. Т. IV, №2, с. 343-354.
51
Павловский О.М. О чем рассказывает обобщенный фотопортрет? // Наука
и жизнь. 1980. № 1, с. 84-90; Юсупов Р.М. О половом диморфизме и
значении женских выборок черепов в антропологии // В сб. Источники
по истории и культуре Башкирии. 1986. Уфа. С. 51-56; Долинова Н.А.
Дерматоглифика удмуртов // В сб. Новые исследования по этногенезу
удмуртов. 1989. Ижевск. Уро АН СССР. С. 108-122.
52
Геодакян В.А. Дифференциальная смертность и норма реакции
мужского и женского пола. Онтогенетическая и филогенетическая пластичность
// Журн. общ. биол. 1974. Т. 35, № 3, с. 376-385.
53
Ehrman L. Nuclear Genes Depending Cytoplasmic Sterility in Dr. paulistorum
// Science. 1964. V. 145, № 3628, p. 159.
54
Геодакян В.А., Геодакян К.В. Новая концепция левшества // Докл.
Акад. Наук. 1997. Т. 356. № 6. С. 1-5.
55
Miyata T., Hayashida H., Kuma K., Mitsuyasu K., Yasunaga T. Male-driven
Molecular Evolution: A Model and Nucleotide Sequence Analysis
// Cold Spring Harbor Symposia on Quantitative Biology. 1987. V.
LII, p. 863-867.
56
Kerkis J. Some Problems of Spontaneous and Induced Mutagenesis
in Mamals and Man // Mutation Res. 1975. V. 29, p. 271-280.
57
Miyata T., Hayashida H., Kuma K., Mitsuyasu K., Yasunaga T. Male-driven
Molecular Evolution: A Model and Nucleotide Sequence Analysis
// Cold Spring Harbor Symposia on Quantitative Biology. 1987. V.
LII, p. 863-867.
58
Shimmin L.S., Chang B. H-J, Li W-H. Male-driven Evolution of DNA
Sequences // Nature. 22 April 1993. V. 362, p. 745-747.
59
Phillips S., Birkenmeier E., Callahan R., Eicher E. Male and Female
Mouse DNAs can be Discriminated using Retroviral Probes // Nature.1982.
V. 297, № 5863, p. 241-243.
60
Brackett B.G., Baranska W., Savicki W., Korovski H. Uptake of Heterologous
Genome by Mammalian Spermatozoa and its Transver to Ova through
Fertilization // Proc. Nat. Acad. Sci. USA. 1971. V. 68. P.
353-357.
61
Кузнецов А.В., Сигаева В.А., Кузнецова И.В., Щит И.Ю. Сперматозоиды
кролика способны связывать чужеродную ДНК // Пробл. репродукции.
1996, № 1. С. 7-10.
62
Grant S., Houben A., Vyskot B., Siroky J., Wei-Hua P., Macas J.,
Genetics of Sex Determination in Flowering Plants // Dev. Genet.
1994. V.15, p. 214-230.
63
см. эпиграф, а также [Геодакян В.А. О теоретической биологии //
В сб. Методологические аспекты эволюционного учения. 1986. Киев.
“Наукова думка”. С. 73-86.]
|